| 爬虫綱・双弓亜綱・主竜類・ 恐竜上目・鳥盤類・鳥脚類・ ハドロサウルス科 Reptilia・Diapsida・Archosauria・ Dinosauria・Ornithischia・ Ornithopoda・Hadrosauridae |
 |
|---|
ハドロサウルス科 (Hadrosauridae)
 |
|---|
| ハドロサウルス科は、中生代白亜紀の北半球に生息した草食恐竜のグループである。 鴨のように長く平たい口吻部が特徴のため、別名、カモノハシ竜、カモハシ竜とも呼 ばれる。発達したクチバシとデンタルバッテリーによって、多様な植物を効率良く餌 とすることができた。幼時期や緊急時は、二足歩行であるが、平常時は、四足歩行で ゆっくりと移動していたことが足跡の化石から推定されている。四肢などの構造は、祖 先であるイグアノドン類に似るが、親指のスパイクは無い。ハドロサウルス科は、系統 としてはイグアノドンや近縁のオウラノサウルスとの関係から白亜紀前期に、ユーラ シア大陸が起源で、出現していたと考えられるが、白亜紀後期に発展し、ヨーロッパや 北米から南米へと広がった。それらは、現在の北極圏内でも発見されている。白亜紀 末期にティラノサウルス科やトリケラトプスとともに絶滅した。K-T(K-Pg)境界のす ぐ下の地層からも化石が発見される数少ないグループの一つである。中空状のとさか を持つランベオサウルス亜科(Lambeosaurinae)と、とさかを持たないか、もしく は、中空でないとさかを持つハドロサウルス亜科(Hadrosaurinae)、サウロロフス 亜科(Saurolophinae)の3つに分けられる。 以下に、ハドロサウルス科の分類と近縁な属を示す。 ハドロサウルス科の分類・近縁属 ハドロサウルス亜科(Hadrosaurinae) ① ハドロサウルス (Hadrosaurus) (北米、白亜紀後期、80–74 Ma) サウロロフス亜科(Saurolophinae) ① エドモントサウルス (Edmontosaurus) (北米・西部、白亜紀後期、73–65.5 Ma) ② クリトサウルス (Kritosaurus) (北米、白亜紀後期、73–66 Ma) ③ グリポサウルス (Gryposaurus) (北米、白亜紀後期、83–74 Ma) ④ サウロロフス (Saurolophus) (北米、モンゴル、白亜紀後期、70–68.5 Ma) ⑤ シャントンゴサウルス (Shantungosaurus) (中国・山東省、白亜紀後期、70 Ma) ⑥ セケルノサウルス (Secernosaurus) (南米・アルゼンチン、白亜紀後期、75 Ma) ⑦ バルスボルディア (Barsboldia) (モンゴル、白亜紀後期、70 Ma) ⑧ ブラキロフォサウルス (Brachylophosaurus) (北米、白亜紀後期、75 Ma) ⑨ プロサウロロフス (Prosaurolophus) (北米、白亜紀後期、76–74 Ma) ⑩ マイアサウラ (Maiasaura) (北米、白亜紀後期、76-74 Ma) ⑪ ウラガサウルス (Wulagasaurus) (中国・黒竜江省、白亜紀後期、65.5 Ma) ⑫ カムイサウルス (Kamuysaurus) (日本・北海道、白亜紀後期、73–66 Ma) ランベオサウルス亜科(Lambeosaurinae) ① アムロサウルス (Amurosaurus) (中国、ロシア、白亜紀後期、66 Ma) ② アラロサウルス (Aralosaurus) (カザフスタン、白亜紀後期、85.8 Ma) ③ オロロティタン (Olorotitan) (ロシア、白亜紀後期、75–67 Ma) ④ カロノサウルス (Charonosaurus) (中国・黒竜江省、白亜紀後期、66.8–66 Ma) ⑤ コリトサウルス (Corythosaurus) (北米、白亜紀後期、77–74 Ma) ⑥ チンタオサウルス (Tsintaosaurus) (中国・山東省、白亜紀後期、70 Ma) ⑦ ニッポノサウルス (Nipponosaurus) (ロシア・サハリン、白亜紀後期、83–80 Ma) ⑧ バクトロサウルス (Bactrosaurus) (中国、白亜紀後期、76.7 Ma) ⑨ パララブドドン (Pararhabdodon) (フランス、スペイン、白亜紀後期) ⑩ パラサウロロフス (Parasaurolophus) (北米、白亜紀後期、76.5–73 Ma) ⑪ ヒパクロサウルス (Hypacrosaurus) (北米、白亜紀後期、75–67 Ma) ⑫ ランベオサウルス (Lambeosaurus) (北米、白亜紀後期、77–75 Ma) ⑬ アングロマスタカトル (Angulomastacator) (北米、白亜紀後期、76.9 Ma) ⑭ ブラシサウルス (Blasisaurus) (スペイン、白亜紀後期、66 Ma) ⑮ ヴェラフロンス (Velafrons) (メキシコ、白亜紀後期、72 Ma) ◎ テルマトサウルス (Telmatosaurus) (ルーマニア、白亜紀後期、70 Ma) |
ハドロサウルス亜科(Hadrosaurinae)
① ハドロサウルス (Hadrosaurus)
 |
|---|
 |
 |
 |
 |
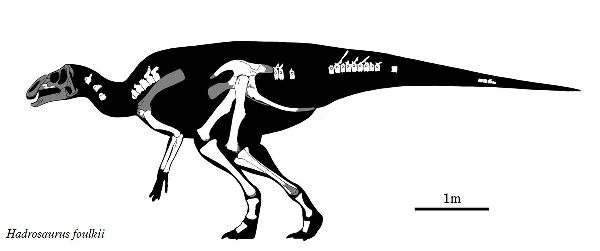
| ハドロサウルスの名は、「頑丈なトカゲ」の意味で名付けられた。化石は、 1838年、ジョン・エスタフ・ホプキンスがニュージャージー州ハドンフィ ールドのクーパー川の小さな支流で大きな骨を発見した。1858年、骨を 見たウィリアム・パーカー・フォルクの興味をそそり、同じ年に白亜紀後期 (約8000万年前)のウッドベリー層から頭蓋骨を欠いた全身の骨格を掘り 出した。フォルクは、古生物学者のジョセフ・ライディと研究し、ハドロ サウルス・ファウルキイと記載し、命名した。この属で唯一の種である。 標本は、世界で最も完全な復元で、展示された史上初の恐竜の骨格になっ た。高さと幅のある胴体を持つ。頭頂部は、とさかを持たず平らである が、鼻づらには、骨質のとさか状の突起がある。吻部には、広がった角質 の嘴を持ち、数百の発達した頬歯がデンタルバッテリーを形成していた。 嘴で摘み取った小枝や葉などをこの歯ですりつぶしていたと考えられる。 後肢は、前肢よりも長く、二足歩行も出来たと考えられるが、大半の時間 を四足歩行で過し、捕食者から逃げる時など緊急時のみ二足になったと される。体長は、約8m、体重は、4tほどだったと推定されている。 |
サウロロフス亜科(Saurolophinae)
① エドモントサウルス (Edmontosaurus)
 |
|---|
 |
 |
 |
 |
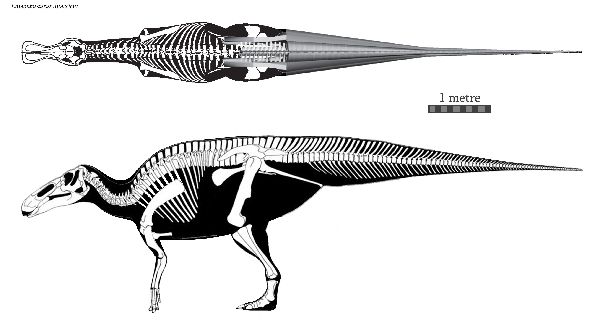
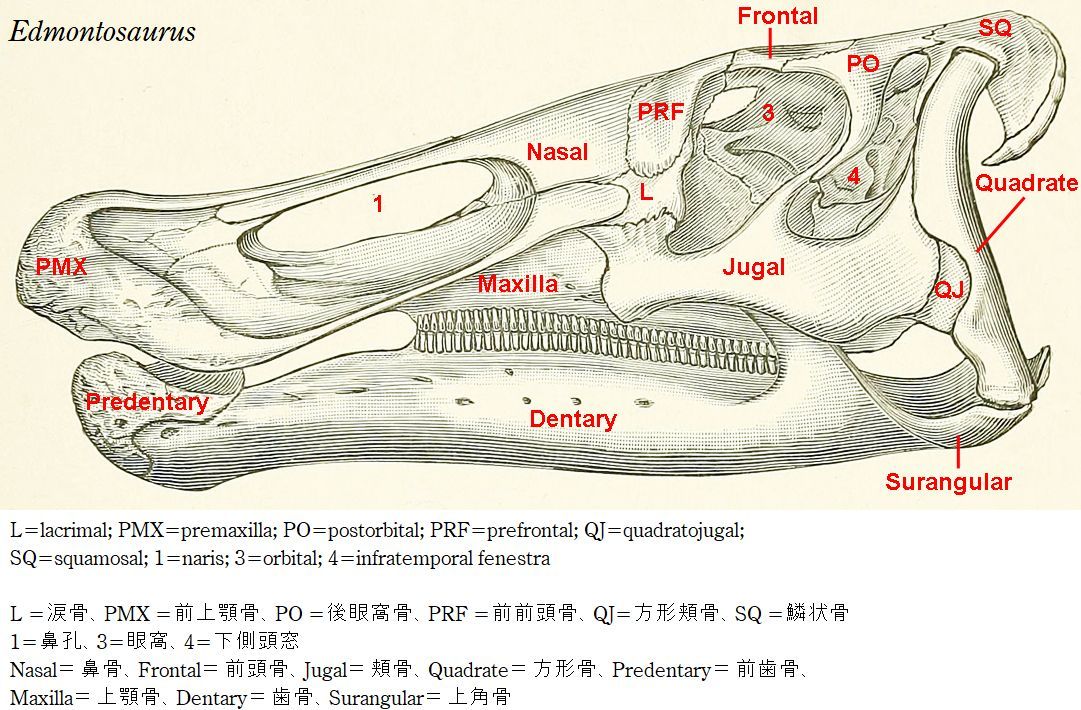
| 最初の発見は、1892年にオスニエル・チャールズ・マーシュによってクラ オサウルス・アネクテンスと名付けられた。後に、E.annectensとされた。 属名の最初は、1917年にローレンス・ランベによってエドモントサウル スと命名された、タイプ種 E. regalisである。この種は、アルバータ州 のホースシューキャニオン累層から発見された。名前は、カナダ、アルバ ータ州南部の首都エドモントンにちなんでいる。エドモントサウルスは、 コロラド州からアラスカの北斜面に至るまで、北アメリカ西部に広く分布 していた。1999年に、「ダコタ」の愛称で呼ばれるミイラが発表された。 ミイラは、皮膚からの残留有機化合物を含んでいる。体のほとんどの領域 が鱗で覆われていたことがわかった。また、エドモントサウルスの頭頂部 に全体が軟骨組織で出来ている「トサカ」があったことがわかった。化 石の分布は、海岸と海岸平野を好んだことを示めしている。エドモントサ ウルスは、2本足と4本足の両方で動くことができる草食動物であった。 ボーンベッドの多量の骨から、エドモントサウルスは、集団で生活して いたと考えられている。全長は、9~13m、体重は、3~4tと推定されて いる。 |
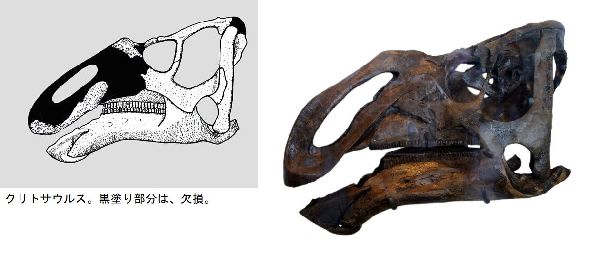
② クリトサウルス (Kritosaurus)
 |
|---|
 |
 |
 |
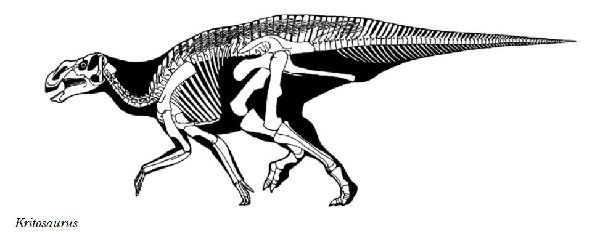
| クリトサウルスは、1904年、アメリカ、ニューメキシコ州、サンファン 郡のオホ・アラモ近郊でバーナム・ブラウンにより、白亜紀後期(7300万 年前)のカートランド累層から発見された。化石は、部分的な頭骨、下顎 骨、いくつかの首から後の骨のみであった。吻部と上側のくちばし(前上 顎骨)の大部分は失われていた。属名は、「分けられたトカゲ」という意味 で、タイプ標本の部分的な頭骨の頬骨の配置にちなんだものである。固有 派生形質(属を識別する特徴)は、前歯骨(下側のくちばし)に鋸歯状の突起 がないこと、下顎のくちばし付近が鋭く下方へ曲がっていること、上顎骨が 重厚で、やや長方形をしていることである。とさかは、完全な状態では鼻 骨から骨のひだのように伸び、両目の間に、後方に折り返すような形で立 ち上がっている。とさかの先端は、ざらざらしている。全長は、約9mほ どと推定される。おそらく、通常、四足歩行の草食恐竜だった。 |
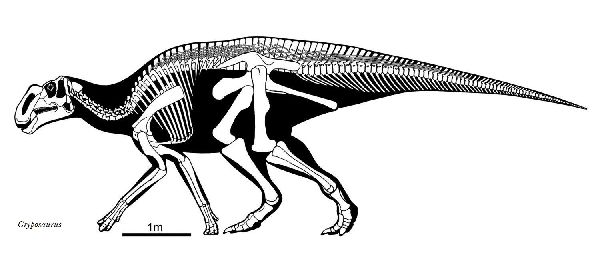
③ グリポサウルス (Gryposaurus)
 |
|---|
 |
 |
 |
| グリポサウルスの名前は、「鷲鼻のトカゲ」を意味する。グリポサウルスの 特徴的な鼻弓(高く緩やかに隆起した鼻骨)は、性別や種の識別や社会的ラ ンク付けなど、さまざまな社会的機能に使用された可能性がある。1913 年にジョージ・F.スタンバーグによって、カナダ・アルバータ州のレッド ディア川沿いの白亜紀後期(7600万年前)のダイナソーパーク累層として 知られている場所から収集された。化石は、頭蓋骨と部分骨格であった。 これは、タイプ種 G.ノタビリスである。グリポサウルスは、クリトサウ ルスに似ており、長年にわたってこの2つは、シノニムであると考えられ ていた。アメリカ・モンタナ州のツーメディスン累層からG.ラティデンス が、ユタ州のカイパロウィッツ累層からG.モニュメンテンシスが知られ ており、テキサス州のジャベリナ累層からも見つかっている。6600万年 前まで生息していた可能性もある。背中の正中線に沿って突き出たピラ ミッド型の鱗など、いくつかの皮膚の印象化石も見つかっている。二足 歩行と四足歩行草食動物であり、川辺に生息していたと考えられてい る。全長は、約8m、体重は、約3tと推定される。 |
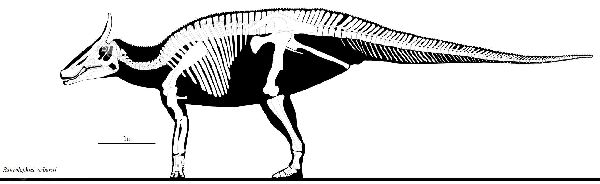
④ サウロロフス (Saurolophus)
 |
|---|
 |
 |
 |
| サウロロフスは、1912年、バーナムブラウンが、アルバータ州のレッド ディア川のトールマンフェリー近くの白亜紀後期(7000万年前)のホース シューキャニオン層から発見した。タイプ種S.osborniと命名した、ほ ぼ完全な骨格であった。全長は、約8.5mと推定される。1946年から19 49年のロシアとモンゴルの古生物学探検隊は、モンゴルのネメグト累層 から数多くの骨格を発見している。S.angustirostris(アングスティロス トリス)と命名された大きな骨格と、さまざまな成長段階の他の骨格も発 見された。全長は、約13m、体重は、約11tと推定される。サウロロフスは、 この2種類で、モンゴルの種は、より長い頭蓋骨(9%)を持ち、鼻の前部 (上顎前骨)は、より上向きであった。また、背中と尾の正中線に沿っ て独特の長方形の鱗の列を持っていた。サウロロフスの最も際立った特 徴は、頭蓋骨の突起である。それは長くてスパイクのようで、目の上か ら始めて、約45°の角度で上に突き出ている。この突起は、鼻骨によっ てほぼ完全に構成されてる。 |
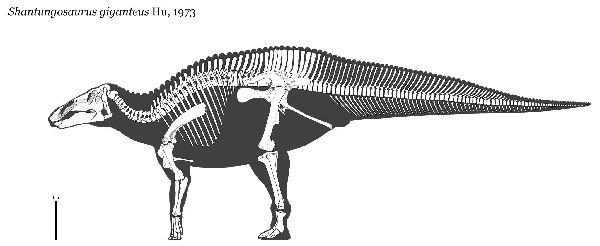
⑤ シャントンゴサウルス (Shantungosaurus)
 |
|---|
 |
 |
 |
| シャントゥンゴサウルスは、「山東トカゲ」を意味する。名前の通り、中 国、山東半島、諸城(Zhucheng)の白亜紀後期の辛格庄(Xingezhuang) 層から発見された。1973年に初めて記載され、5つ以上の不完全な骨格 が知られている。シャントゥンゴサウルスは、最大の鳥盤類の一つであり、 ホロタイプの頭骨は、長さ1.63mで、北京の中国地質学研究所に展示さ れる復元骨格では、体長約15mである。最大、体長17m、体重16tに達 したと推測されている。大型化の特長として前足が、他のハドロサウル ス類よりも長く発達している。これは、巨体を支えるためほとんど4本 足で生活するための適応と考えられる。エドモントサウルスと非常によ く似ている。しかし、一般にはその巨大さと、尾の骨の特徴が異なり、 クチバシは、横に大きく広がり、下顎の骨が厚くなっていることなどか ら、別の恐竜と考えられている。 |
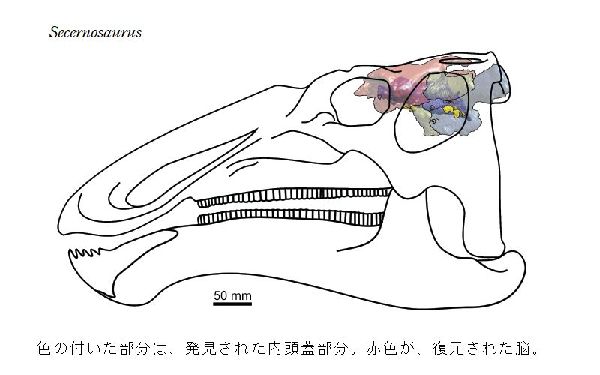
⑥ セケルノサウルス (Secernosaurus)
 |
|---|
 |
 |
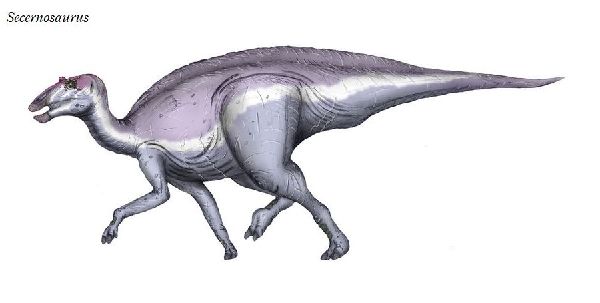 |
| セケルノサウルスは、南アメリカで1923年に発見され、1979年に命名 された数少ないハドロサウルス類の最初のものである。1923年にJ・B・ アボット率いるフィールド博物館の遠征の際、アルゼンチンのチュブ州、 パタゴニア中南部にある白亜紀前期(7100~6600万年前)のコルウエ・ ウアピ湖累層から発見された。頭蓋および後頭蓋などの部分骨格である。 ユーラシア大陸と北アメリカが一体となったラウラシア大陸に住んでいた ほとんどのハドロサウルスとは異なり、南アメリカに住んでいた。セケル ノサウルスの祖先は、白亜紀後期に南北アメリカの間に一時的に陸橋が形 成され、2つの大陸間の移動が可能なときに、南アメリカに渡ったことが 示唆されている。よって、「分離した爬虫類」を意味する、セケルノサウ ルスと名付けられた。クリトサウルスの近縁種である。タイプ標本の体長 は、約5mほどだったと推定されている。幼体かもしれない。ハドロサウ ルス科としては、小さかった。 |
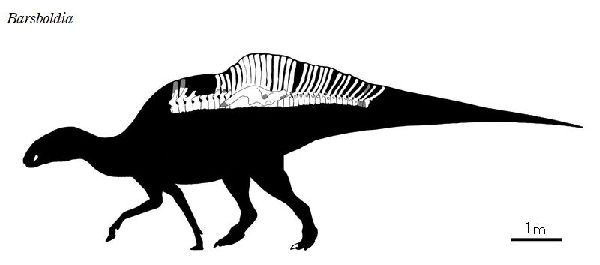
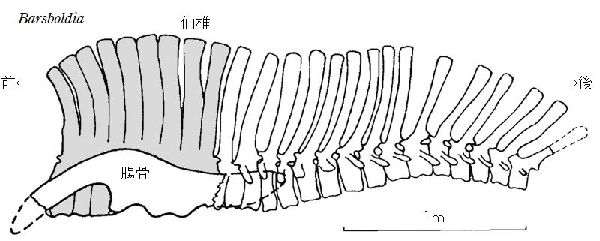
⑦ バルスボルディア (Barsboldia)
 |
|---|
 |
 |
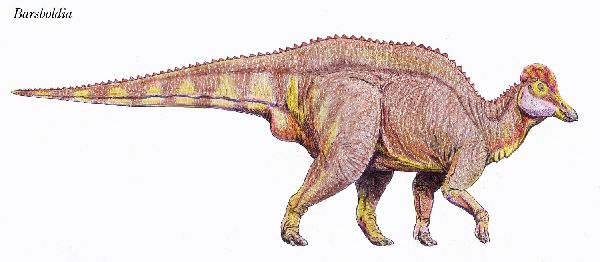 |
| バルスボルディアの名は、「バルスボルド(著名なモンゴルの古生物学者) のもの」という意味がある。化石は、1970年、ポーランド・モンゴルの遠征 隊により、モンゴル、ウムヌゴビ県オムノギョフの白亜紀後期(約7000万 年前)のネメグト累層から発見された。発見された骨格は、頭骨や肢骨等を 欠いていた。9個の胴椎、9個の仙椎、15個の尾椎、左側の腸骨及び左右 の恥骨の1部分、7個の肋骨、後肢の断片が大まかに脊椎に関節した状態 であった。この骨格の最も顕著な特徴は、神経棘で、特に仙椎で高く、ヒ パクロサウルス・アルティスピヌスに次ぐ高さであり、最初の数個の尾椎 では、神経棘の先端が棍棒形である。頭部が発見されていないものの、は じめは、仙骨の底部のキールからランベオサウルス亜科の特徴とされた、 しかし腸骨の特徴からサウロロフス亜科に分類された。骨格は、ヒパク ロサウルスのものに良く似ている。体長は、12~14m、体重は、5tほどだ ったと推定されている。 |
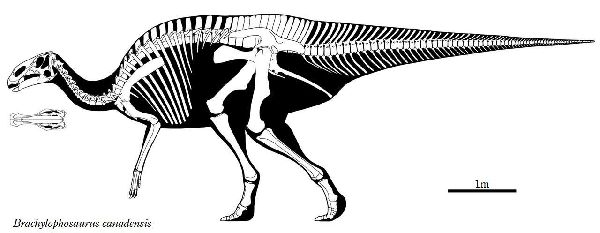
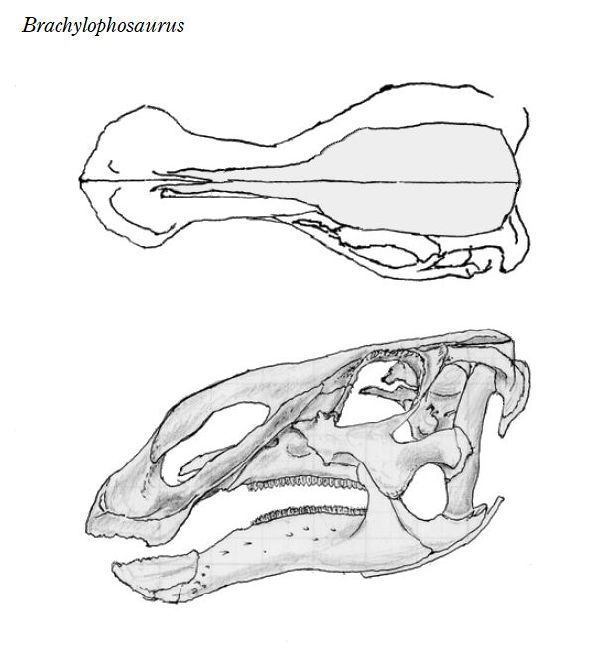
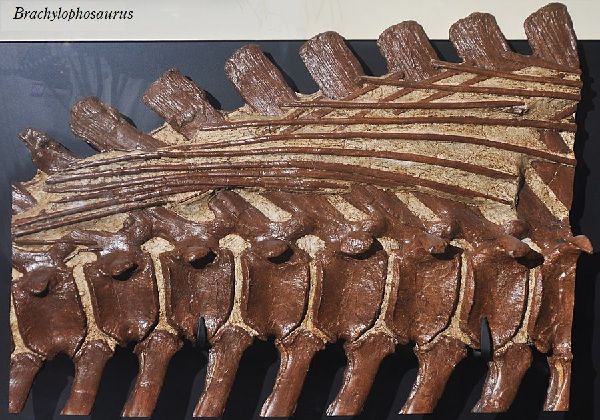
⑧ ブラキロフォサウルス (Brachylophosaurus)
 |
|---|
 |
 |
 |
 |
 |
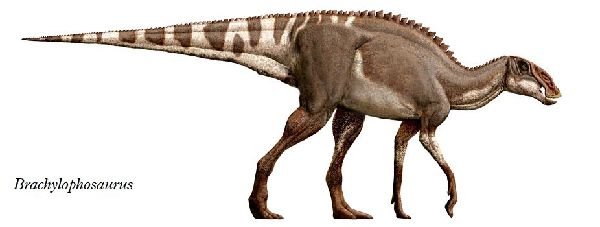 |
| ブラキロフォサウルスは、「短いトサカ付きのトカゲ」を意味する名前で ある。1936年にカナダ、アルバータ州、スティーブビル近郊の白亜紀後 期(約7700万年前)のオールドマン累層で頭蓋骨と部分骨格が発見され、 1953年にチャールズ・スタンバーグにより記載され、グリポサウルスと された。1988年にジャック・ホーナーがモンタナ州のジュディスリバー 層で発見された化石をブラキロフォサウルス・グッドウィニ(Brachylo- phosaurus goodwini)と命名し、アルバータ州のものも同種とされた。 1994年、モンタナ州フィリップス郡のマルタで、アマチュア古生物学者 のネイト・マーフィーは、彼が「エルビス」と名付けた完全で粉砕されて いないブラキロフォサウルスの骨格を発見した。2000年、亜成体のブラ キロフォサウルスの完全に関節し、部分的に「ミイラ化した」骨格であ る「レオナルド」が、ダン・スティーブンソンによって発見された。最も 壮観な恐竜の発見の1つと見なされている。胃の内容物からシダ、苔類、 淡水藻類、針葉樹、モクレン等の被子植物の葉や枝など40種類以上の植 物を食べていたことがわかった。ブラキロフォサウルスの特徴は、頭蓋 骨の上部が成長し、張り出した平らな舌形の大きな骨板を形成する。頂 上は、正中線上に低い縦方向の尾根を持っている。前肢が長く、おそら く主に四足歩行であった。体長は、9~11m、体重は、7tほどだったと推 定されている。 |
⑨ プロサウロロフス (Prosaurolophus)
 |
|---|
 |
 |
 |
 |
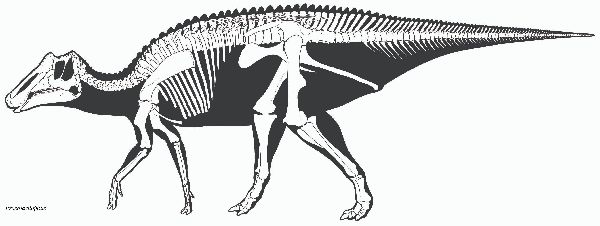
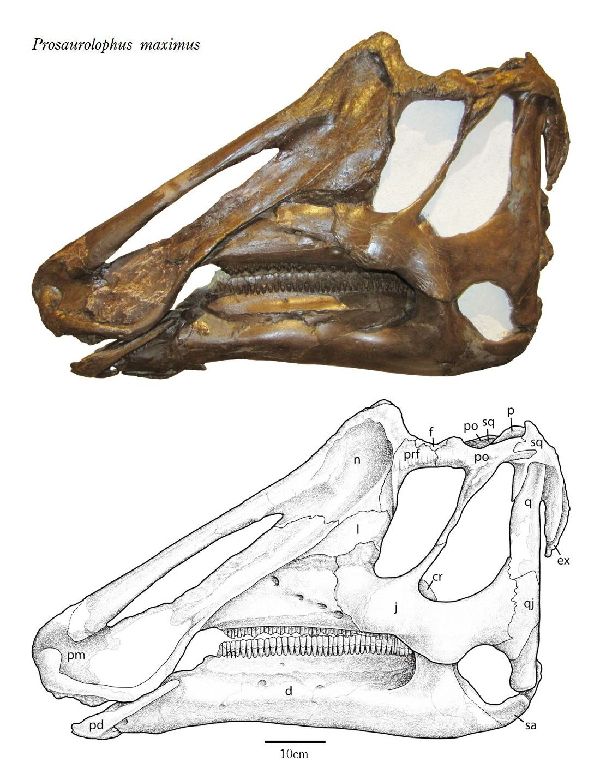
| プロサウロロフスのタイプ種は、P.maximusで、1916年にアメリカ自然 史博物館のバーナム・ブラウンによって記載された。第2の種、P.black- feetensisは、1992年にロッキー博物館のジャック・ホーナーによって 記載された。2種は、主にとさかの大きさと頭骨の比率によって識別され る。化石は、1915年、カナダ、アルバータ州、スティーブビル近くのレッ ドディア川の白亜紀後期(7600万年前~7500万年前)のダイナソーパー ク累層(Dinosaur Park Formation)で頭蓋骨と顎が発見された。おお よそ同じ時代のアメリカ、モンタナ州、グレーシャー郡のツーメディスン 累層(Two Medicine Formation)のボーンベッドからも多量に発見さ れている。目の前に頑丈で短い三角形のとさかを持っている。とさかの 側面は、凹んでくぼみを形成している。側面は、P.blackfeetensisでは P.maximusより勾配があって高かった。少なくとも、P.blackfeetensis では、とさかは、成長にともなって目に対して後方へと移行した。上腕は、 比較的短かった。体長は、約8.5m、体重は、3tほどだったと推定されて いる。 |
⑩ マイアサウラ (Maiasaura)
 |
|---|
 |
 |
 |
 |
 |
 |
 |
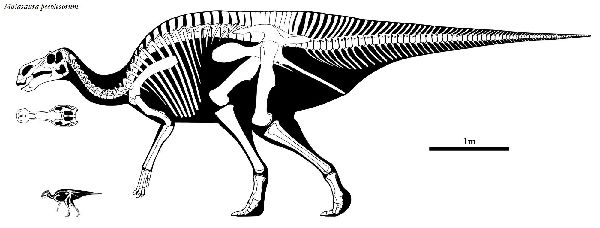
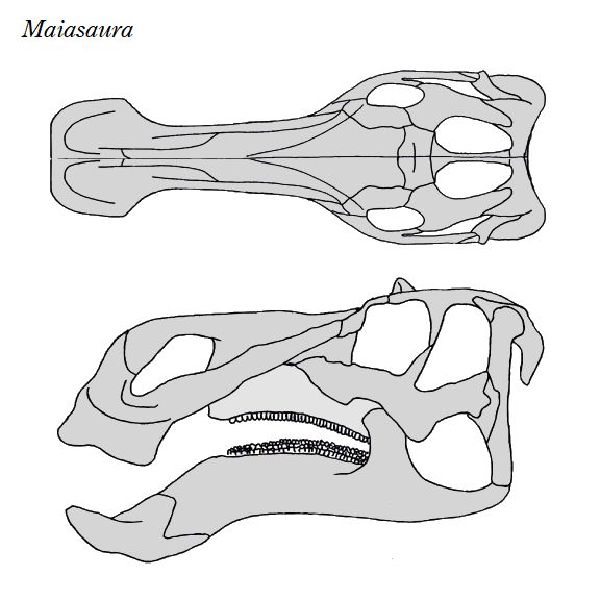
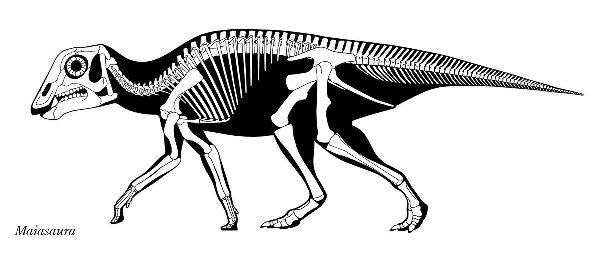
| マイアサウラは、「良い母親トカゲ」という意味の属名がつけられた。化 石は、1979年にモンタナ州バイナム在住のローリー・トレクスラーによ って、モンタナ州西部のチョトー近くの白亜紀後期(約8360万年前~約 7210万年前)のツーメディスン累層から頭蓋骨が発見された。眉間に低 いトサカがある。恐竜古生物学者ジャック・ホーナーとロバート・マケラ によって新種のホロタイプと認定され、タイプ種をマイアサウラ・ピー ブルソラム(Maiasaura peeblesorum)と名付けた。この土地でジョン とジェームズ・ピーブルズの家族は、1978年に直径約2mのすり鉢状の 巣と、巣の中や外に15体の幼体の化石を発見した。これらの発見は、他 の発見につながり、この地域は、営巣コロニーで「エッグマウンテン」 として知られるようになった。巣と巣の間は、約7mで、成体の長さより 短い。巣は、土でできており、円形または、らせん状のパターンで産卵さ れたダチョウの卵くらいの大きさの30〜40個の卵が含まれていた。卵は 親が巣に入れた腐った植生に起因する熱によって孵化した。孵化すると、 赤ちゃんマイアサウラは、彼らの足が完全に発達していなかったため、歩 くことができなかったことを示している。化石は、また、彼らの歯が部分 的に摩耗していたことを示している。それは、大人が巣に食物を持ってき たことを意味する。これは、巨大な恐竜が、子育てと餌を与えた最初の 証拠だった。マイアサウラは、主に幼体として二足歩行であり、年をと るにつれてより四足歩行の姿勢に切り替えることもわかった。体長 は、8~9m、体重は、4tほどだったと推定されている。 |
⑪ ウラガサウルス (Wulagasaurus)
 |
|---|
 |
 |
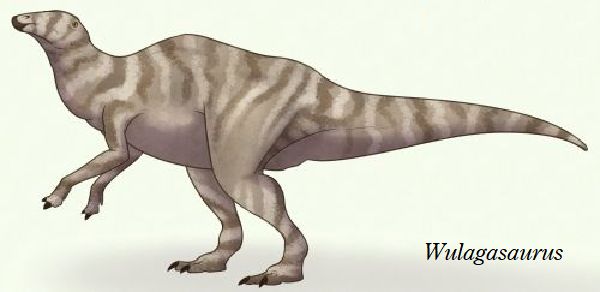 |
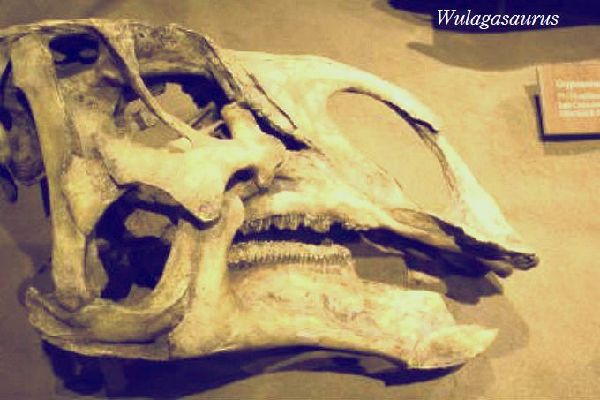
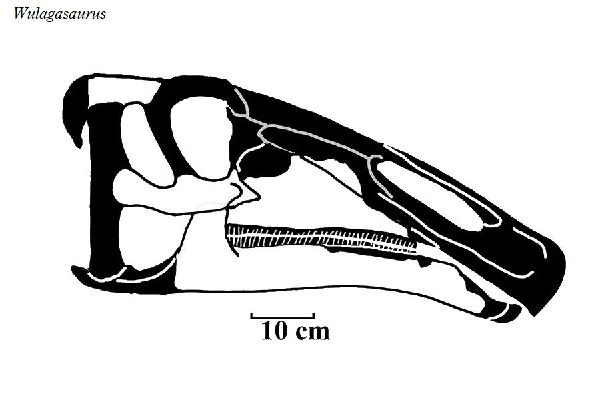
| ウラガサウルスは、2000年以降、中国・黒竜江省、アムール川地域、ウ ーラガ採石場の白亜紀後期(6900万年前~6600万年前)のユリアンゼ累 層(Yuliangze Formation)の骨床で発見された。化石は、部分的な歯骨 である。その後、脳蓋3個、頬骨1個、上顎骨2個、歯骨、肩甲骨2個、胸 骨2個、上腕骨2個、坐骨1個が発見された。上腕の独特の形と細い歯から、 ハドロサウルス科と判断され、2008年に、ベルギーの古生物学者のパス カル・ゴドフロワ(Pascal Godefroit)らによって、ウラガサウルス・ドン ギ(Wulagasaurus dongi)と命名された。北米のブラキロフォサウルス およびマイアサウラと多くの形態学的類似点を共有している。基底的な 特徴から、すべてのハドロサウルス科がアジア起源であることを示唆して いると考えられている。通常は、二足歩行、食事の時などは、四足歩行で あった。体長は、約9m、体重は、3tほどだったと推定されている。 |
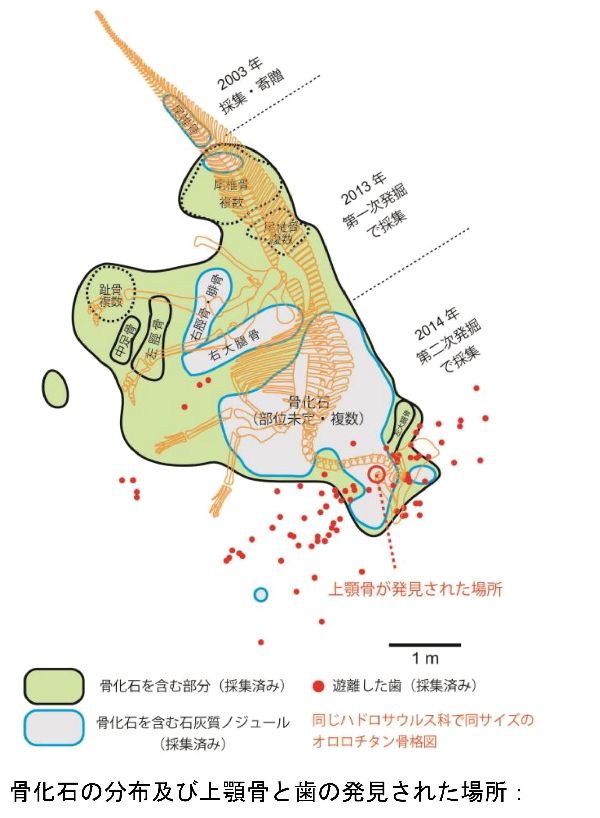
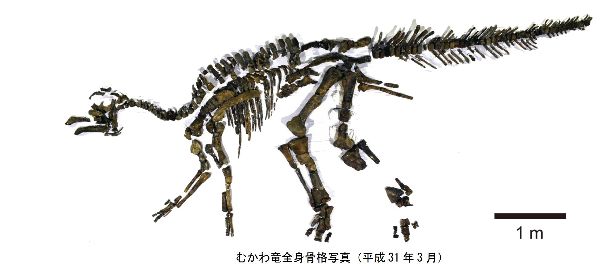
⑫ カムイサウルス (Kamuysaurus)
 |
|---|
 |
 |
 |
 |
 |
 |
 |
 |
| カムイサウルスは、2003年、北海道、穂別町(当時)在住の堀田良幸によ って、むかわ町穂別地区の白亜紀後期(7200万年前)の外側陸棚堆積物の 海成層、蝦夷層群函淵層から連結した13個の尾椎骨が発見された。はじ め、首長竜とされ、穂別博物館に収蔵されていた。2010年から2011年に、 東京学芸大学所属の佐藤たまき准教授が首長竜の化石を調査していて、 穂別博物館を訪れ、クリーニングをしてみて、首長竜ではなく、恐竜の 骨の特徴を持つことを明らかにした。同年、北海道大学総合博物館の小 林快次准教授に鑑定を依頼し、ハドロサウルス科の尾椎骨であると同定 していた。発掘された地層は、波の影響を受けないほど水深の深い場所 で堆積したこともあり、全身骨格が埋蔵している可能性が高いと判断し、 第1次(2013年9月から同年10月)と第2次(2014年9月)発掘を実施した。 2018年3月には、ほぼすべての化石のクリーニング作業が完了し、その 結果、この化石が骨の個数を分母にすると60%、総体積を分母にすると 80%にも及ぶ完成度のきわめて高い全身骨格化石であることが分かった。 鼻、仙椎の一部、指骨だけが欠けていた。研究を進めたところ、エドモ ントサウルス類に属し、ロシアのケルベロサウルスと中国のライヤンゴ サウルスにも近縁な恐竜であることがわかった。ブラキロフォサウルス の亜成体が持つトサカのような、薄く平たいトサカがあったと思われる。 その他、3つの固有の特徴と、13のユニークな特徴を併せ持っているこ とがわかり、新属新種であることが判明した。2019年、カムイサウルス ・ジャポニクス(Kamuysaurus japonicus)と命名された。通称は、「 むかわ竜」ということになった。日本で恐竜の全身骨格化石が発掘され たのは、樺太で発掘されたニッポノサウルスを含めれば、福井県勝山市で 発掘されたフクイベナートルと合わせて3例目となる。かつて海であっ た地層から発掘されたハドロサウルス科の全身骨格化石としても世界で 3例目であった。前肢が華奢なので、通常は二足歩行、食事の時などは 四足歩行であった。体長は、約8m、体重は、4~5.3tほどだったと推定 されている。 |
ランベオサウルス亜科(Lambeosaurinae)
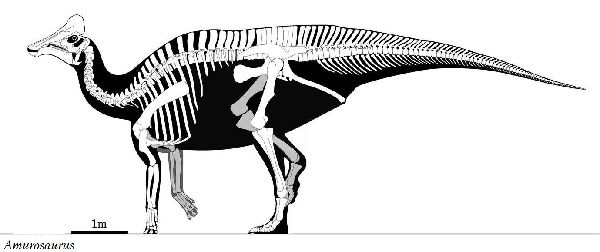
① アムロサウルス (Amurosaurus)
 |
|---|
 |
 |
 |
 |
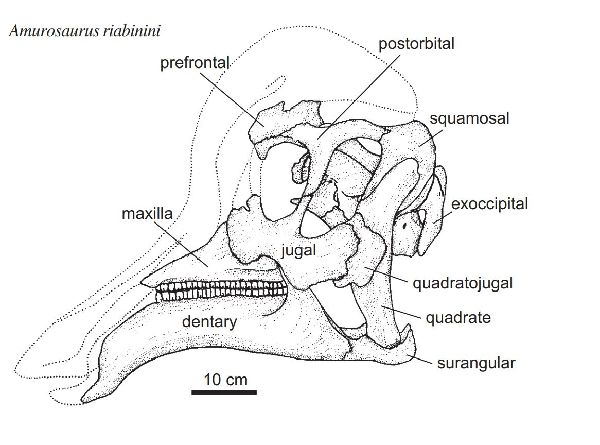
| アムロサウルスは、1984年、ロシアのアムール州ブラゴヴェシチェンス クの白亜紀後期(6600万年前)のUdurchukan層で発見されており、ロシ ア東部と中国北東部に分布する Tsagayan 層群において最古の層である。 この地層は、ボーンベッドを形成している。堆積物は、化石をわずかな 距離だけ運搬して川の氾濫原に横たえたとされる。ボーンベッドの露出 している領域は、広くないが、これまで発見された化石の90%は、大半 は幼体であるランベオサウルス亜科のアムロサウルスに属し、残りは、 サウロロフス亜科に属するケルベロサウルスである。獣脚類の歯も豊富 で、捕食動物や腐肉食動物に由来する傷が骨に数多く確認されている。 ロシアの古生物学者ユーリ・ロボトスキーとセルゲイ・クルザノフが、 1991年に初めてアムロサウルスを記載・命名した。ホロタイプ化石は、 同一個体の左半身に由来する単一の上顎骨と歯骨のみからなる。しかし、 夥しい数の別個体ではあるものの、頭骨の他の骨や全身の骨の大半がボ ーンベッドに保存されている。鶏冠は、発見されていないものの、大半 のランベオサウルス亜科と同様に、アヒルの嘴のような形状の吻部と内 部が空洞の鶏冠を持つ二足歩行の植物食性動物だったとみられている。 成体の化石は、希少である。属名は、発見地のアムール川にちなんでい る。通常は、二足歩行、食事の時などは四足歩行であった。体長は、約 8m、体重は、3tほどだったと推定されている。 |
② アラロサウルス (Aralosaurus)
 |
|---|
 |
 |
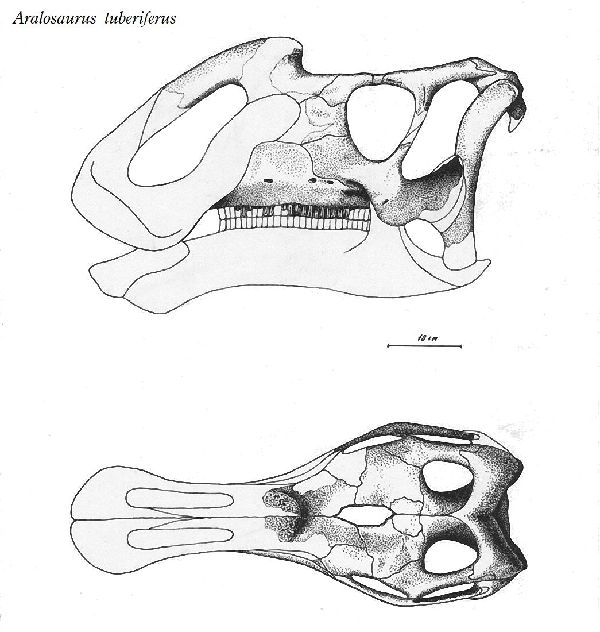
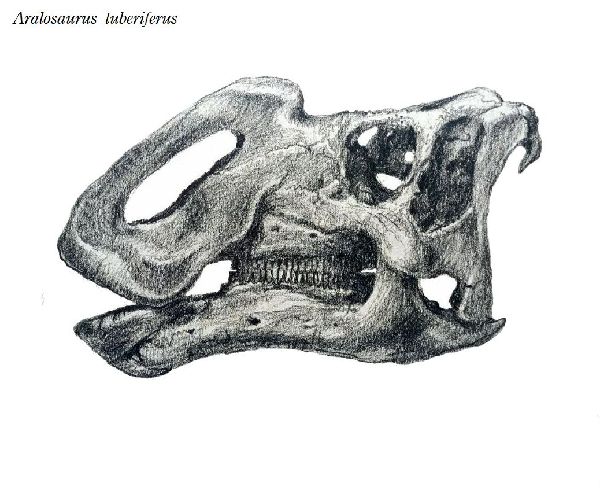
| アラロサウルスは、1957年、カザフスタン中部、シャフシャフ地域の近 くの白亜紀後期(8580万年前)のボストーベ累層(Bostobe Formation) から産出した。属名は、発見地近くのアラル海にちなんでいる。化石は、 頭蓋骨(鼻の大部分と下顎骨全体を欠いている)、孤立した歯と尺骨、橈 骨、大腿骨、脛骨、腓骨、距骨、中足骨などであった。1968年にアナト リー・コンスタンチノヴィッチ・ロジェストヴェンスキーによって記載・ 命名されたアラロサウルス・ツベリフェルス(Aralosaurus tuberifer- us)という1つの種のみが知られている。鼻骨の後部が、眼窩の前で急激 に上昇するため、塊茎を持っているという意味の種名が与えられた。こ の鼻の塊茎は、明らかに中空で、呼吸器系の一部を囲み、鼻腔が拡張し ている。典型的なランベオサウルスの特徴である。通常は、二足歩行、 食事の時などは、四足歩行であった。体長は、約6.5m、体重は、4tほど だったと推定されている。 |
③ オロロティタン (Olorotitan)
 |
|---|
 |
 |
 |
 |
 |
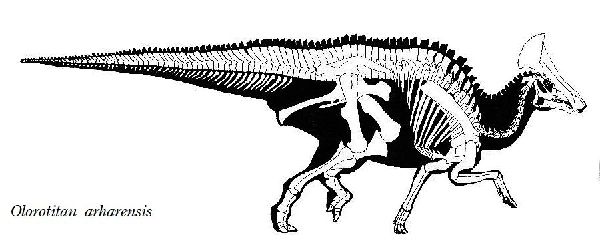
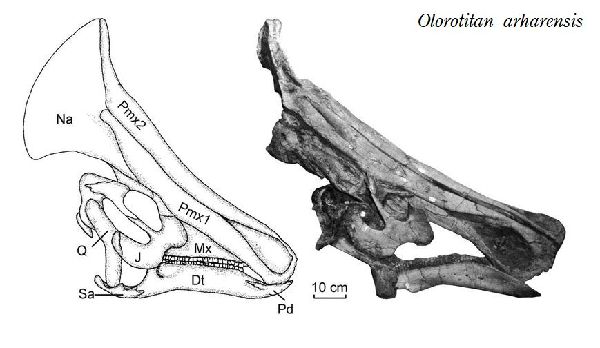
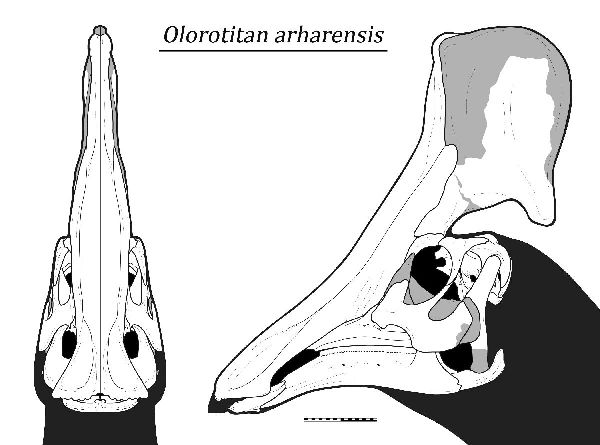
| オロロティタンは、1999年から2001年にかけて、ロシア東部アムール 地区クンドゥルの白亜紀後期(7200万年前)のウドゥルチュカン層(Ud- urchukan Formation)から産出した。化石は、ほぼ完全な骨格で、2003 年に唯一の種であるオロロティタン・アルハレンシス(Olorotitan arha- rensis)と記載・命名された。属名は、「巨大なハクチョウ」を意味する。 これまで、北米以外で発見された最も完全なランベオサウルス類の骨格で ある。後方に向いた扇形、または斧型の鶏冠状突起によって特徴づけら れる。高く広い中空の鶏冠状突起は、鼻腔と繋がっており、おそらく視 覚と聴覚によって同種を識別できるように機能したと思われる。頭骨は 18個の頸椎で構成された長い首に支えられている。それまでに知られて いたハドロサウルス類の頸椎は、最大でも15個であった。仙骨も15~ 16個の椎骨で成るが、少なくとも他のハドロサウルス類よりも3個多 い。通常は二足歩行、食事の時などは四足歩行であった。白亜紀-古第 三紀の絶滅イベント中に絶滅した。最後の恐竜の1つである。体長は、 約8m、体重は、2.6~3.4tほどだったと推定されている。 |
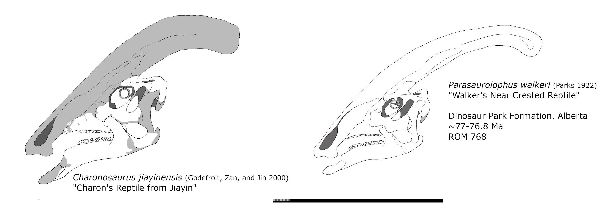
④ カロノサウルス (Charonosaurus)
 |
|---|
 |
 |
 |
 |
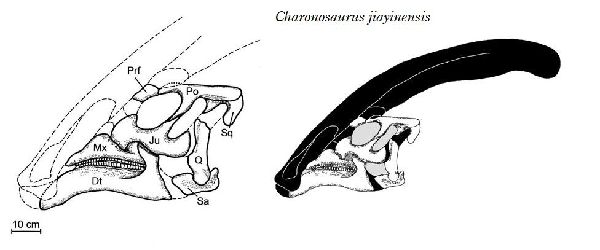
| カロノサウルスは、1975年以降、中国北東部の黒竜江省嘉陰村の西にあ る白亜紀後期(6600万年前)のユリアンゼ累層(Yuliangze Formation) から産出した。ホロタイプ標本は、断片的な頭蓋骨である。2000年にパ スカル・ゴドフロワ、シュチン・ザン、リヨン・ジンによって、カロノサ ウルス・ジアイネンシス(Charonosaurus jiayinensis)として記載・命 名された。属名は、ギリシャ神話に登場するアケロン川の渡し守カロー ンにちなんでいる。ランベオサウルス類は、北米では、7200万年前頃に は姿を消したが、アジアでは、白亜紀の終わりまで生き残ったことを示 している。地層は、数十平方メートルに及ぶボーンベッドを形成し、比 較的強い流れのある河川環境で形成され、恐竜の死骸が当時の低い地点 に集中して積み上げられたことを示している。化石の約90%が、ランベ オサウルスの骨で、残りは、アンキロサウルス、獣脚類、カメ、ワニのも のである。肉食恐竜の歯の豊富さは、カロノサウルスの死骸が引き裂か れて捕食者やスカベンジャーによって消費されたことを示している。部 分的な頭蓋骨は、パラサウロロフスの頭蓋骨に似て、長く後方に突き出 た中空のトサカを持っていた。カロノサウルスは、現在アジアで知られ ている最大のハドロサウルスの1つである。通常は四足歩行であった。 体長は、約10~13m、体重は、5tほどだったと推定されている。 |
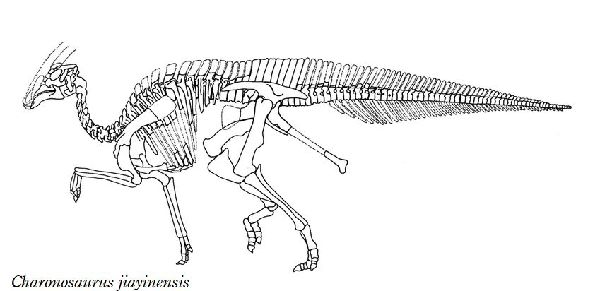
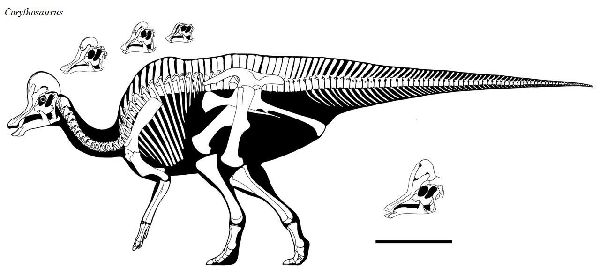
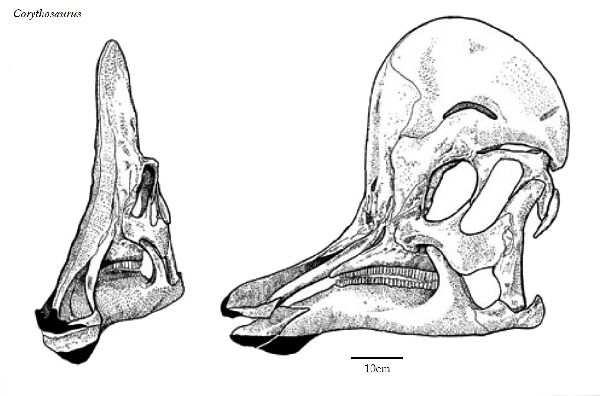
⑤ コリトサウルス (Corythosaurus)
 |
|---|
 |
 |
 |
 |
 |
 |
| コリトサウルスは、1911年に、カナダ、アルバータ州のレッドディア川 の白亜紀後期(7750万年前~7480万年前)のベリーリバー層群(Belly River Formation)よりバーナム・ブラウンによって発見された、ほぼ完 全なホロタイプを含む多くの標本から知られている。1914年にバーナム ・ブラウンによって、コリトサウルス・カジュアリウス(Corythosaurus casuarius)と命名された。名は、「コリント式のトカゲ」を意味し、そ の頭部の形態が、古代ギリシアの一部族コリント族の兜にある頭飾りに 似ているため、これにちなんだ属名にした。名の由来ともなった半円状 の突起が、頭頂部に突き出している。この突起は、性差が見られ、雄の 方が大きかったと推定されている。この突起は、鼻骨が拡張したもので あり、鼻道が入り組んでいるため、外観と違って内部の構造は、相当複 雑となっている。この突起は、鳴き声の増大などに寄与したと考えられ ている。脊椎は、棘突起が伸長し、背面に稜を形成していた。尾椎は、 胸胴椎から続く骨質の結合組織に固められ、可動性は乏しかった。ホロ タイプ標本は、尾の最後の部分と前脚の一部だけが欠けていたが、多角 形の鱗の印象も保存されていた。通常は、二足歩行、食事の時などは、 四足歩行であった。体長は、約10~13m、体重は、4~5tほどだったと 推定されている。 |
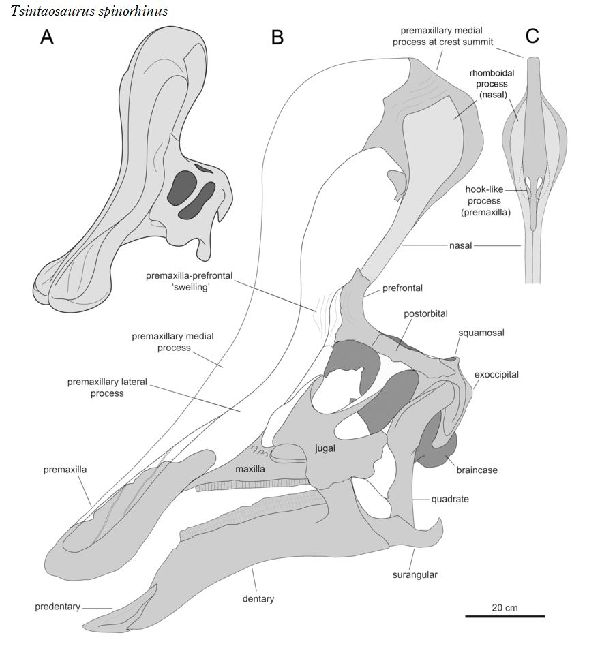
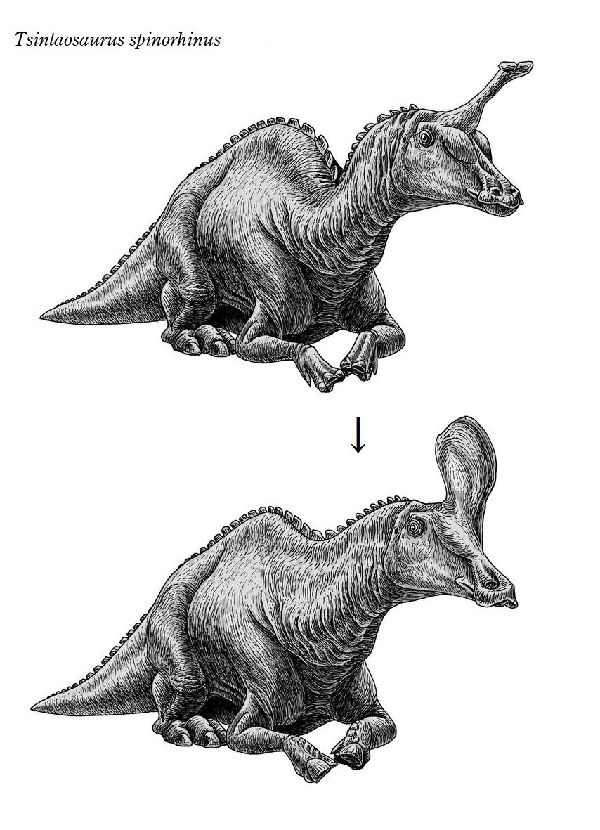
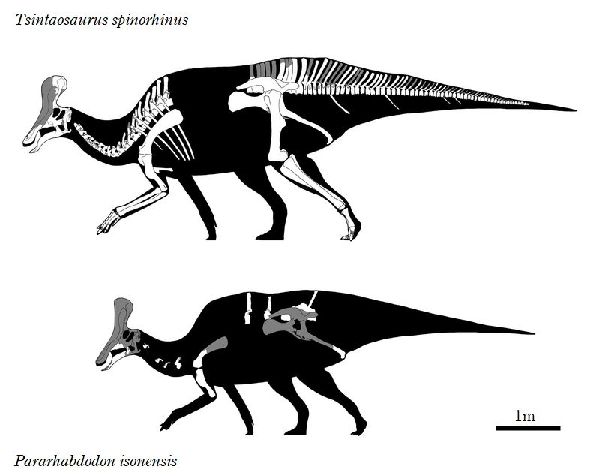
⑥ チンタオサウルス (Tsintaosaurus)
 |
|---|
 |
 |
 |
 |
 |
 |
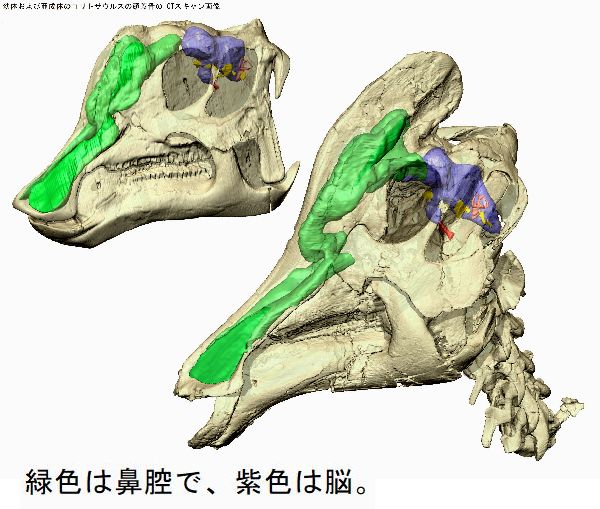
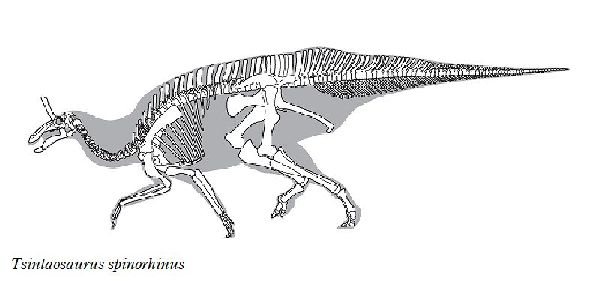
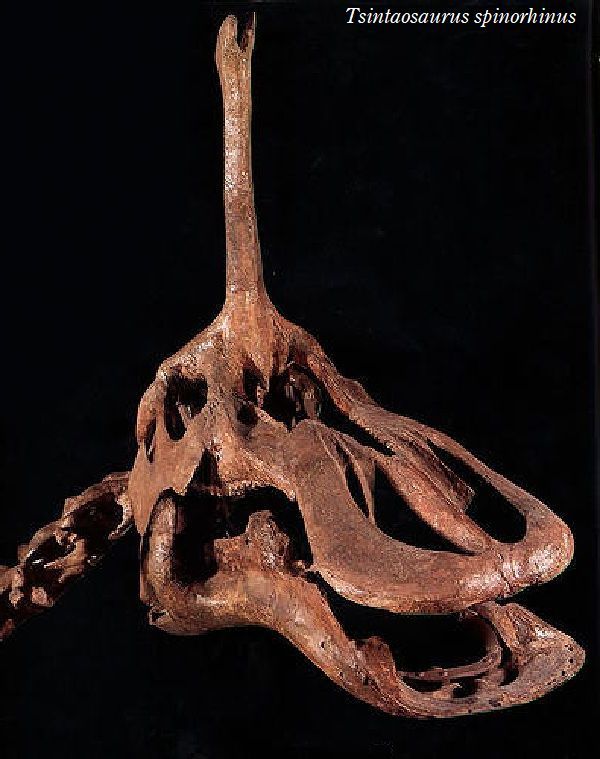
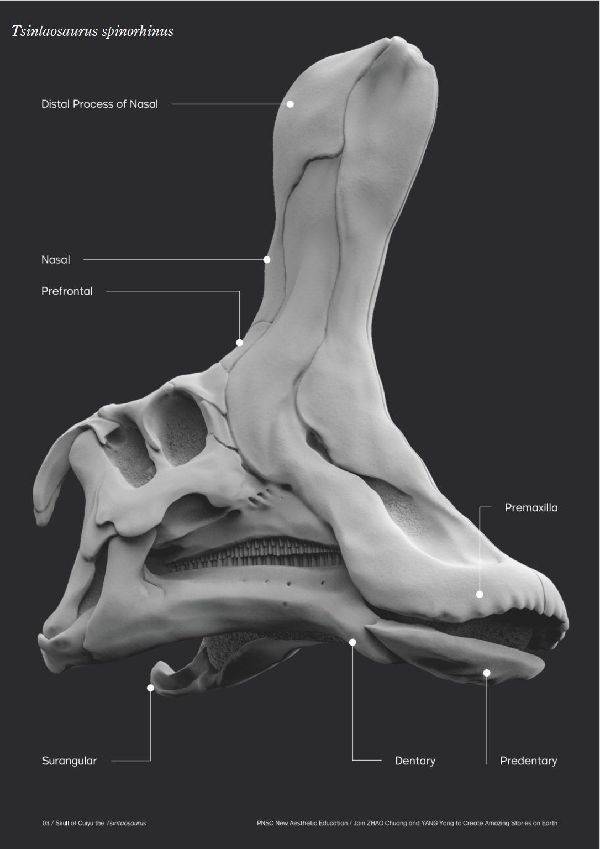
| チンタオサウルスは、1950年、中国東部山東省青島市近くの萊陽市の呂 格荘金剛口村北西、冲溝の道路側にある白亜紀後期(7000万年前)のジン ガンコウ累層で古生物学者、周明鎮とその教え子たちが大型のハドロサ ウルス類の脛骨と腓骨を発見した。1958年に中国の古生物学者C.C.ヤン グによって最初に記述され、チンタオサウルス・スピノルヒヌス(Tsin- taosaurus spinorhinus)と命名された。発掘された頭蓋骨は、細長い 突起が突き出た奇妙な頭骨であった。2013年、ドイツ・バイエルン州立 古生物学・地質学博物館のアルバート・マルケス(Albert Prieto-Márq- uez)とテキサス州立大のジョナサン・R・ワーグナー(Jonathan R.W- agner)が、「生前のチンタオサウルスの頭部のトサカは、もっと大きく てドーム状になっていた」という仮説を発表した。化石では、たまたま、 ドーム部分の後ろ半分しか残っていなかった。通常は、二足歩行、食事 の時などは四足歩行であった。体長は、約10m、体重は、3tほどだったと 推定されている。 |
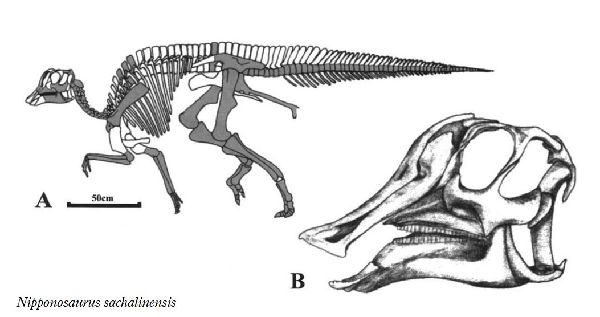
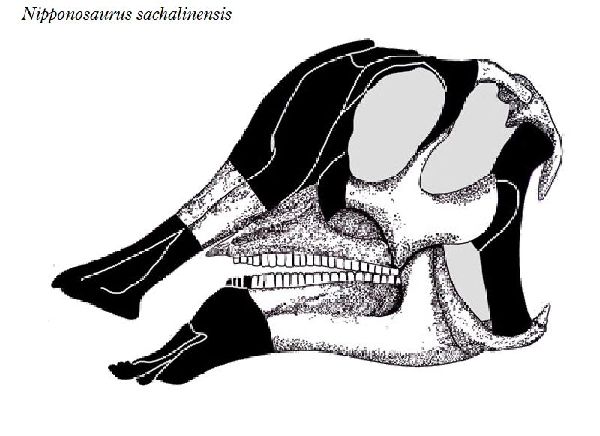
⑦ ニッポノサウルス (Nipponosaurus)
 |
|---|
 |
 |
 |
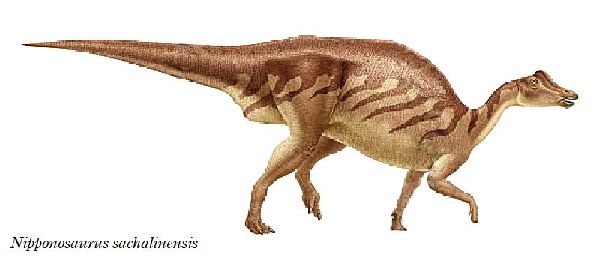 |
| ニッポノサウルスは、1934年(昭和9年)に、樺太庁豊栄郡川上村の三井 鉱山、川上炭鉱施設内の病院建設現場(現、ロシア連邦サハリン州ユー ジノ・サハリンスク)で、根本要氏や中西義男氏などによって、白亜紀後 期(8000万年前)の竜ヶ瀬層から発見された。化石は、頭骨の一部、骨盤、 腰椎、後脚など、全身の約40%が産出している。産出層は、白亜紀に形 成された海成層であり、死後、死体が海に運ばれたか、海岸で死亡した と考えられている。日本人によって研究、記載されたはじめての恐竜で ある。1936年、北海道帝国大学、長尾巧教授が、ニッポノサウルス・サ ハリネンシス(Nipponosaurus sachalinensis)と命名した。1937年、 再度同じ場所で調査が行われ、手足の化石が発掘された。泥岩の地層か らは、パラプゾシア・ジャポニカとスフェノセラムス・シュミッティと 呼ばれる化石軟体動物が発見されている。この恐竜は、ヒパクロサウル スに近縁と考えられている。歯のないクチバシがあり、頭頂にはドーム 状の小さな膨らみ(トサカ)がある。通常は二足歩行、食事の時などは四 足歩行であった。体長は、約4m、体重は、1tほどだったと推定されてい るが、幼体であったと考えられている。 |
⑧ バクトロサウルス (Bactrosaurus)
 |
|---|
 |
 |
 |
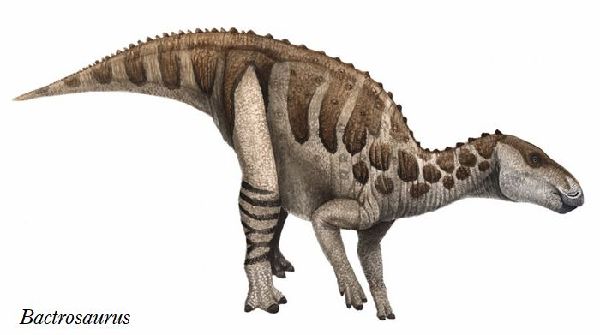 |
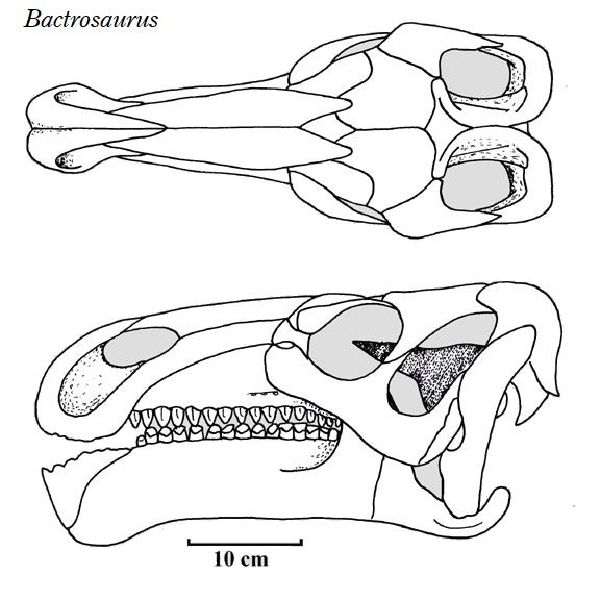
| バクトロサウルスは、1922年~1923年に、アメリカ自然史博物館のロ イ・チャップマン・アンドリュースの中央アジア探検隊が、中国、内モ ンゴル自治区、二連浩特市のゴビ砂漠の白亜紀後期(約9000万年前)のイ レンダバス層(Iren Dabasu Formation)から発見した。1933年アメ リカの古生物学者チャールズ・ギルモア(Charles W. Gilmore)が、新 属新種としてバクトロサウルス・ジョンソニ(Bactrosaurus johnsoni) を記載・命名した。属名は、脊椎から棍棒(baktron)のような大きな神経 棘が突き出ていることから名付けられた。最初のバクトロサウルスの化 石は、1個体の B.johnsoniの部分骨格で、頭蓋骨と骨盤、脚の骨で構 成されていた。収集された標本は、孵化したばかりの個体から成体まで、 さまざまな年齢層のものであった。外見上は、イグアノドンに似ている が、初期のカモノハシ竜の特徴を持っている。ハドロサウルス科すべて の属の祖先と考えられている。一つの歯槽に3本の歯が縦に並んでいる (イグアノドン類は縦に2本)。親指のスパイクが無い。通常は二足歩 行、食事の時などは四足歩行であった。体長は、約6m、体重は、1.2tほ どだったと推定されている。 |
⑨ パララブドドン (Pararhabdodon)
 |
|---|
 |
 |
| パララブドドンは、1980年代半ばに、スペイン、リェイダ、イソナ近く のピレネー山脈のサントロマダベラの白亜紀後期(6600万年前)のトレン プ累層から発見された。トレンプ累層は、河口からデルタの環境を特徴 とする大陸からわずかに海洋の河川湖沼環境に堆積した。化石は、当初 イグアノドン類のラブドドン属に割り当てられたが、比較的遠縁である ことがわかった。1993年に、カサノバ・クラデラス(Casanovas Clad- ellasらは、新種パララブドドン・イソネンシス(Pararhabdodon iso- nensis)と名付けた。1999年に、ランベオサウルス亜科に属することが 示された。2009年には、確かにランベオサウルスであり、より具体的に は、中国のチンタオサウルス属の近縁種であるという証拠が提唱された。 化石は、頭蓋後のさまざまな部位、主に椎骨、および頭蓋骨の上顎が含 まれる。尾椎と仙椎の神経棘は、長く、他のハドロサウルス類同様に高 く盛り上がった背中をしていた。白亜紀-古第三紀の絶滅イベント中に 絶滅したと考えられ、化石記録から知られている最後の非鳥類恐竜の1つ である。通常は二足歩行、食事の時などは四足歩行であった。体長は、 約5m、体重は、500Kgほどだったと推定されている。 |
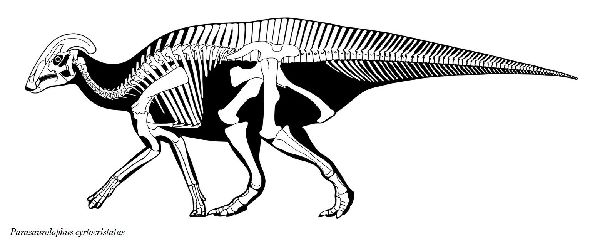
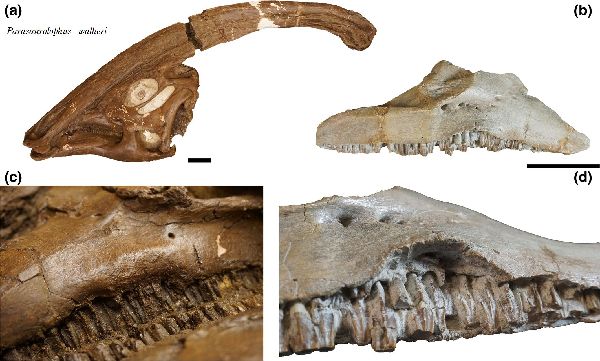
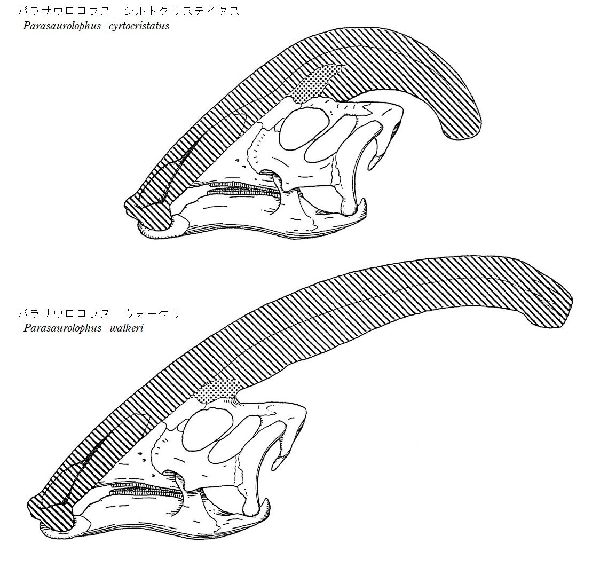
⑩ パラサウロロフス (Parasaurolophus)
 |
|---|
 |
 |
 |
 |
 |
| パラサウロロフスは、1920年、トロント大学のフィールドパーティーが、 カナダ、アルバータ州のレッドディア川沿いのサンドクリークの近くの 白亜紀後期(7650万年前)の恐竜公園累層から産出した。発見された化石 は、尾と後肢のほとんどが欠けた、頭蓋骨と部分的な骨格であった。19 22年、カナダのウィリアム・アーサー・パークス(William Arthur Parks) は、最初に発見されたタイプ標本をパラサウロロフス・ウォーケリ(Para- saurolophus walkeri)と命名した。その後、モンタナ州のヘルクリーク 累層やニューメキシコ州サンファン郡のカートランド累層からも頭蓋骨 が発見された。最も顕著な特徴は、頭の後ろから突き出た、上顎前骨と 鼻骨で構成されていた頭蓋稜(トサカ)である。このトサカは、中空で、各 鼻孔からトサカの端まで通じる個別の管があり、方向を逆にして頂上を 下って頭蓋骨に戻った。このトサカの湾曲の程度や長さ、形で種類が分 けられている。雌雄の差もある。腕は、ハドロサウルス科としては比較 的短く、短いが広い肩甲骨を持っていた。後肢や骨盤は、他のハドロサ ウルス科と比較すると頑丈であった。通常は二足歩行、食事の時などは 四足歩行であった。体長は、10~15m、体重は、4~18tほどだったと推 定されている。 |
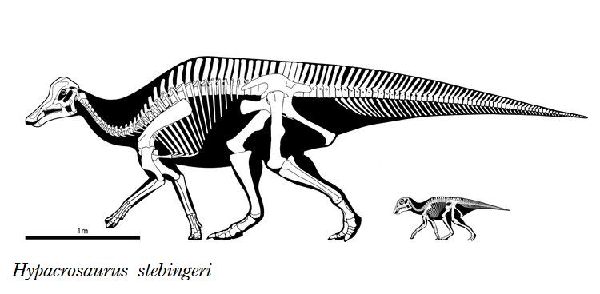
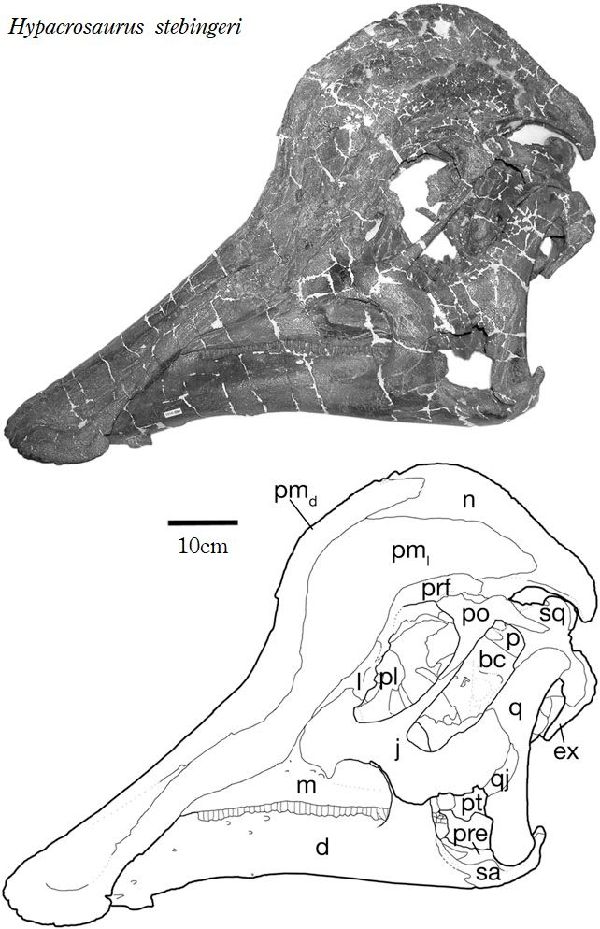
⑪ ヒパクロサウルス (Hypacrosaurus)
 |
|---|
 |
 |
 |
 |
 |
 |
| ヒパクロサウルスは、1910年、バーナム・ブラウンが、カナダ、アルバ ータ州、Tolman Ferry近郊のレッドディア川沿いの白亜紀後期(7500 万年前)のホースシュー・キャニオン累層で発見した。標本は、アメリカ 自然史博物館のために収集された。この化石は、いくつかの椎骨と部分 的な骨盤を含む頭部を含まない部分骨格であった。すぐに、2つの頭骨 が発見され、記載された。1913年にバーナム・ブラウンが、ヒパクロサ ウルス(Hypacrosaurus)と命名した。この属は、H.altispinusとH.st- ebingeriの2種が知られ、少なくとも北米の化石の多い中空のトサカを 持つカモノハシ恐竜としては、最新の属であった。ヒパクロサウルスの 空洞のトサカは、個体が性別や種を識別できるようにする視覚信号や、 音を立てるための共鳴室を提供するなどの社会的機能を持っていた可能 性が高い。ランベオサウルス亜科の中でもコリトサウルスとオロロティ タンに近縁である。高い神経棘とトサカによって他の中空のトサカを持 つランベオサウルス亜科と最も容易に識別できる。この神経棘は、背中 では各々の椎骨の椎体の5~7倍の高さがある。H.stebingeriは、モン タナ州レイシャー郡のツーメディスン累層の頂上近くで発見された、 卵と巣を持つ孵化したばかりの幼体を含むさまざまな化石にちなんで名 付けられた。18.5~20cmのほぼ球形の卵を生み、中には体長60cmほ どの胚が入っていた。孵化したばかりの幼体は、体長1.7mほどであった。 胚や幼若な個体は、のちにトサカを形成するわずかな骨の膨らみがある 頭骨を持っていた。2~3年で性的成熟し、10~12年で最大サイズに達 したと思われる。通常は二足歩行、食事の時などは四足歩行であ った。体長は、約9.1m、体重は、4tほどだったと推定されている。 |
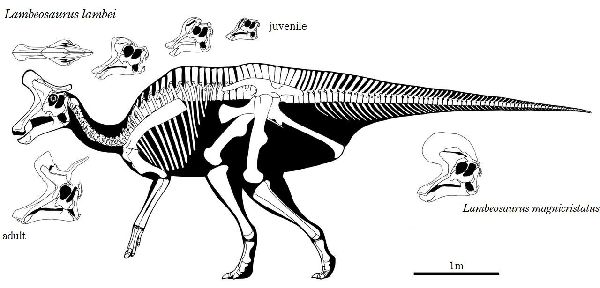
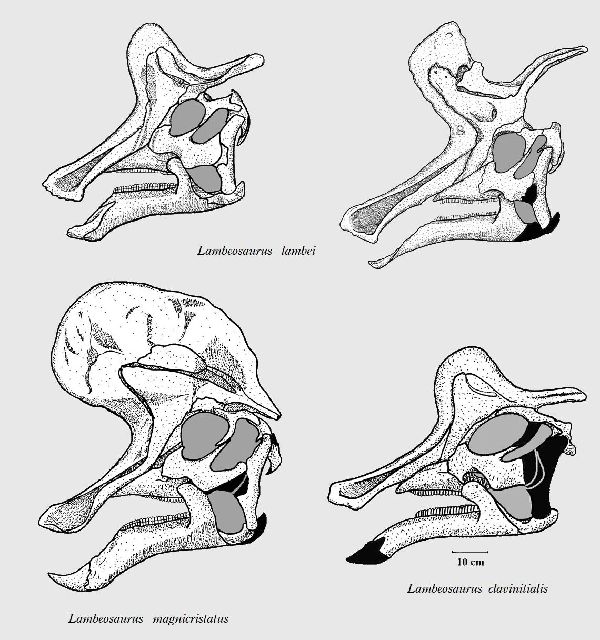
⑫ ランベオサウルス (Lambeosaurus)
 |
|---|
 |
 |
 |
 |
 |
| ランベオサウルスは、1902年にローレンス・ランベがアルバータ州のハ ドロサウルス科の四肢と他の骨を発見したことに始まる。1923年、ウィ リアム・パークス(William Parks)は、カナダ、アルバータ州の白亜紀後 期(7600万年前)の恐竜公園累層から発見した頭蓋骨に、ランベオサウル ス ランベイ(Lambeosaurus lambei)と、命名した。1919年に亡くな った、ローレンス・ランベを称えて名付けた。最も有名な特徴は、ミトン に似ていた独特の中空の頭蓋骨のトサカである。ランベオサウルスの場合 トサカは、コリトサウルスと似ているがやや小さく、前傾している。ま た頭部後方に向かって緻密骨の棒が一本突き出し、容易に区別がつく。 トサカの形状に関しては、全体的に小ぶりであったり、もしくは後部の 骨の突起がなかったりと、個体間でやや差がある。かつては、これを元 に個々に別の種であると考える研究者もいた。しかし現在ではこれは性 別、あるいは成長の違いによるものだと考えられている。化石は、アル バータ州のほか、バハ・カリフォルニア半島とモンタナ州でも発見され てい。足跡化石からは、群れをつくり、四足歩行で移動していたこと がわかっている。体長は、約9m、体重は、1~3.5tほどだったと推定さ れている。 |
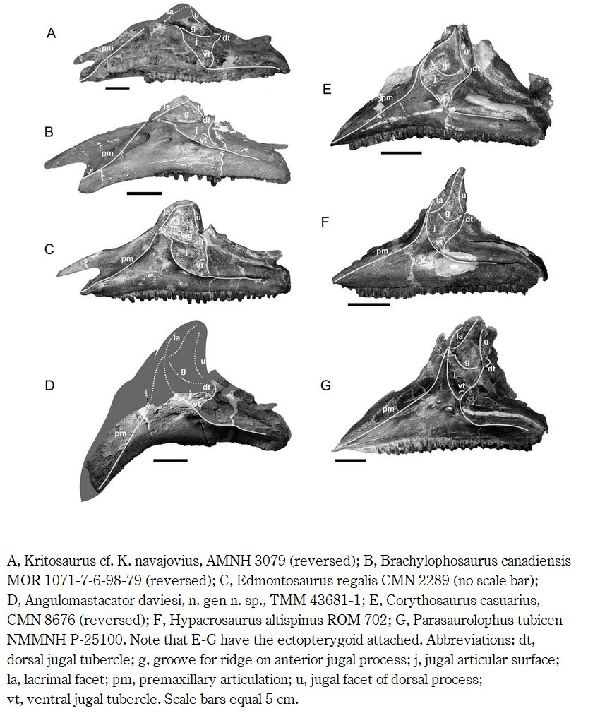
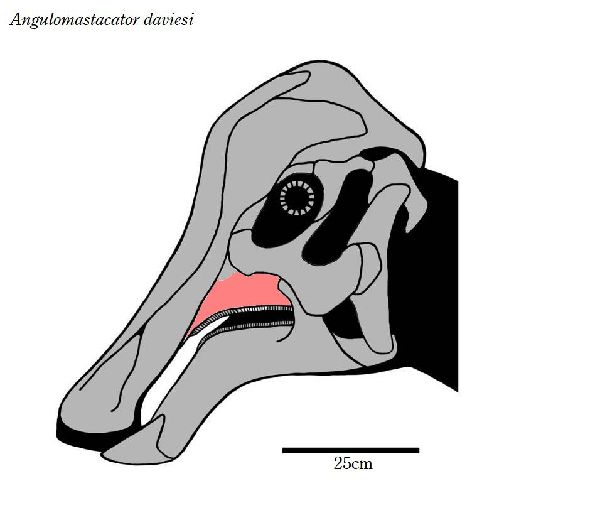
⑬ アングロマスタカトル (Angulomastacator)
 |
|---|
 |
 |
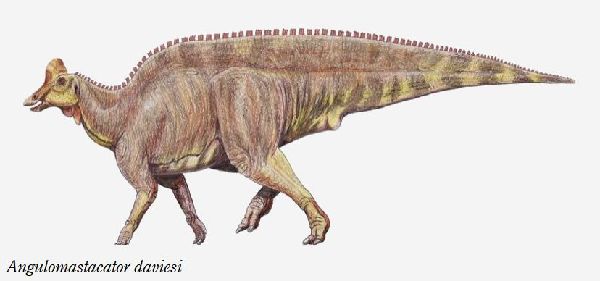 |
| 1983年、カイル・L・デイビスが、テキサス州リオグランデ、ビッグベ ンド国立公園の白亜紀後期(7690万年前)のアグジャ累層(Aguja Form- ation)から中空のクレストを持つランベオサウルスの仲間が産出したこ とを発表した。2009年にワグナー(Wagner)とレーマン(Lehman)に よって記載され、アングロマスタカトル・ダヴィエシ(Angulomasta- cator daviesi)と命名された。ホロタイプの化石は、唯一の部分的な左上 顎骨である。この骨は、他のハドロサウルスとは異なり、前端が約45°下 方に湾曲しており、歯列もそこに収まるように曲がっている。他の地層 からは報告されていない上顎の独特な特徴は、アグジャ層の恐竜が、種 としての終末期の形態であったという仮説を支持するものである。通常 は二足歩行、食事の時などは四足歩行であった。体長・体重は、化石が 不足しているので不明である。 |
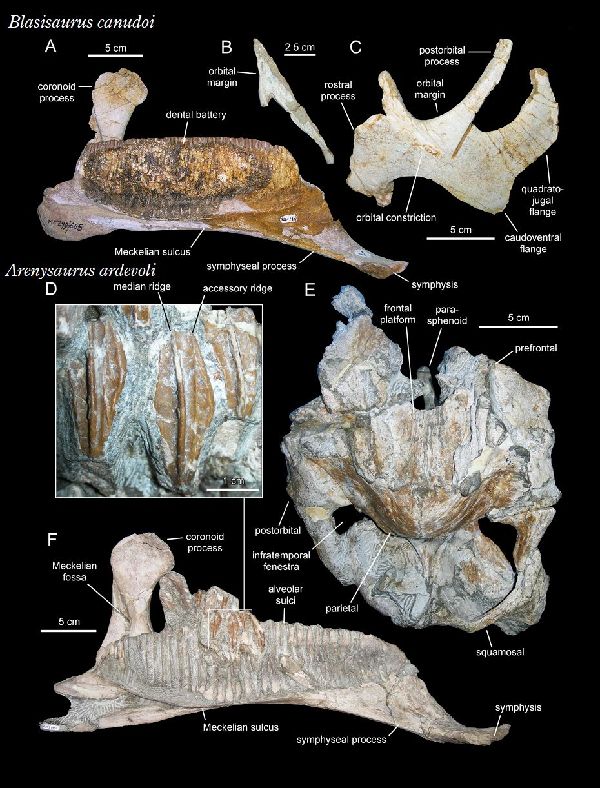
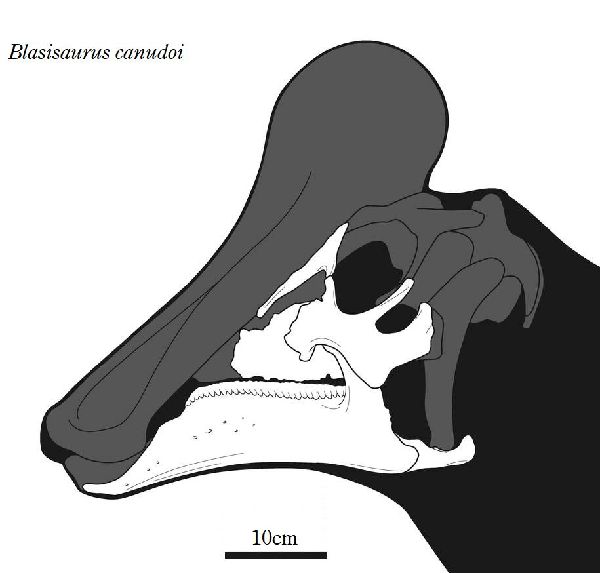
⑭ ブラシサウルス (Blasisaurus)
 |
|---|
 |
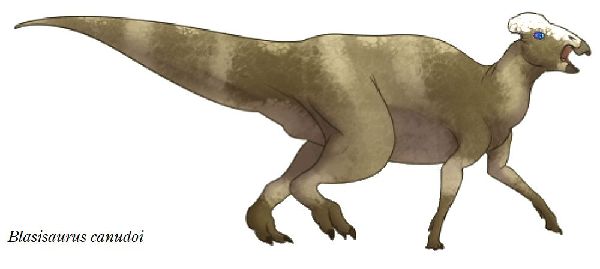 |
| ブラシサウルスは、1989年、スペイン、アラゴン州ウエスカ県、ブラシ 1地域(Blasi 1)の白亜紀後期(6600万年前)のアレン層(Arén Format- ion)から産出した。ホロタイプ標本は、断片的な下顎がついた頭骨からな る。2010年にスペインのペネロペ・クルザド・カバレロ(Penélope Cr- uzado Caballero)らの研究者チームによりブラシサウルス・カヌドイ( Blasisaurus canudoi)と命名された。ホロタイプ標本は、左頬骨であ る。パラタイプ標本は、左上顎骨の2つの断片、右涙骨、左歯骨、右上角骨 である。ブラシサウルスの特徴は、頬骨がフック状の上端を持つ後部突 起を持っていることである。同一の層からは、近縁なアレニサウルスが 発見されているが、ブラシサウルスの歯表面の二次隆起が存在しないこ とや歯の形状からも識別がなされている。ブラシサウルスは、下顎の前 縁が下方へ湾曲している点でパララブドドンとも異なる。通常は二足歩 行、食事の時などは四足歩行であった。体長・体重は、化石が不足してい るので不明であるが、下顎サイズから、体長は、約4.2m、体重は、360 Kgほどだったと推定されている。 |
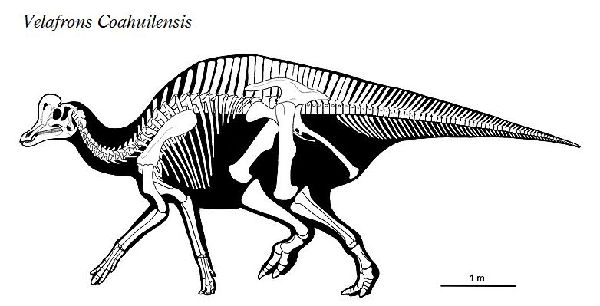
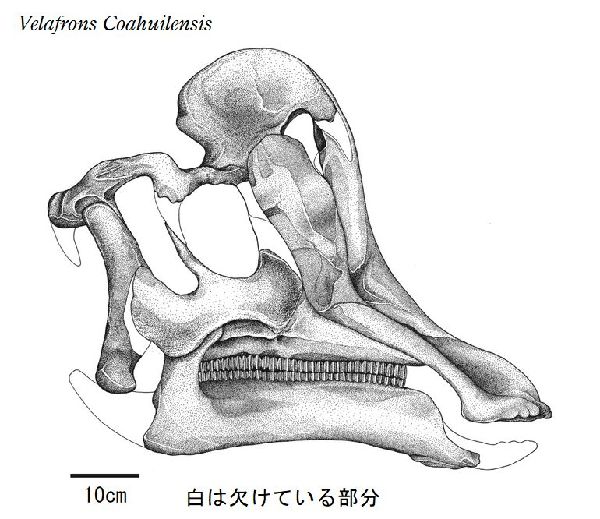
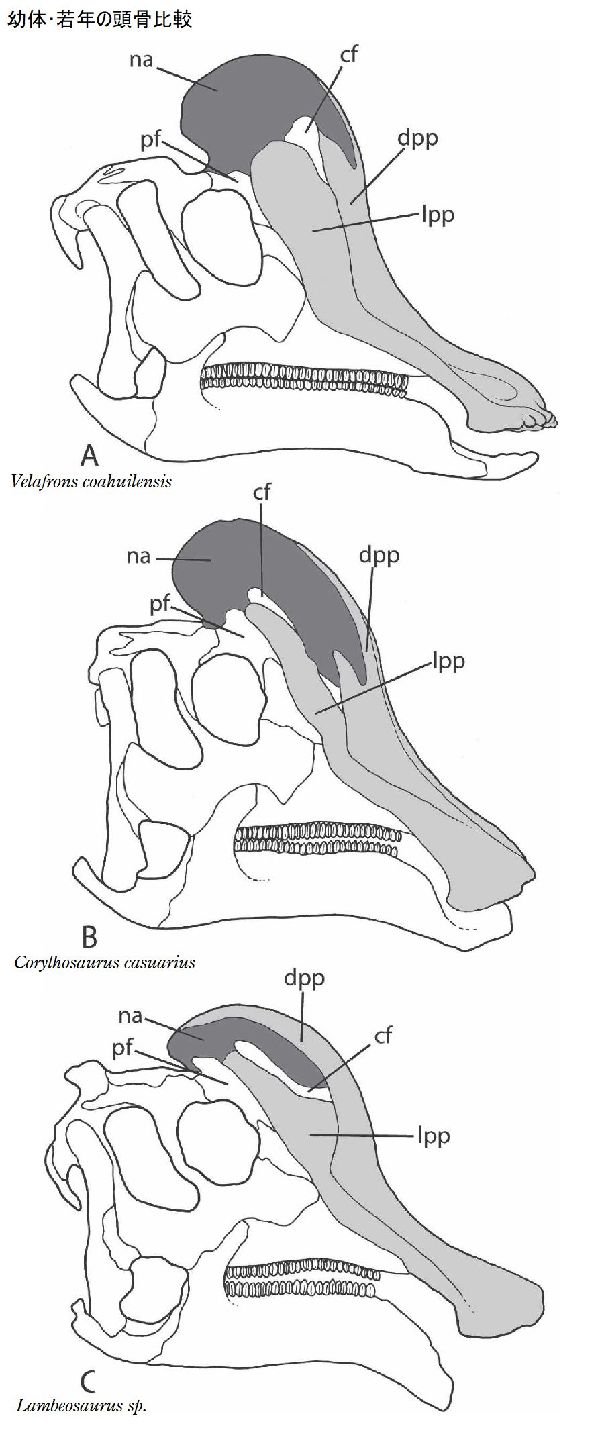
⑮ ヴェラフロンス (Velafrons)
 |
|---|
 |
 |
 |
 |
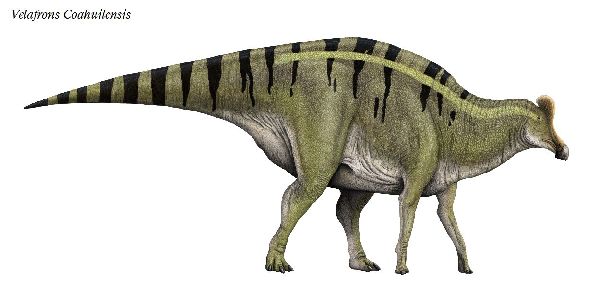 |
| ヴェラフロンスは、1995年に発見・発掘が開始され、2002年、メキシ コ、コアウイラ州リンコンコロラド近郊の白亜紀後期(7350万年前)のセ ロ·デル·プエブロ累層(Cerro del Pueblo Formation)から発掘が完了 した。2007年、テリー・A・ゲイツ(Terry A.Gates)らによって記載さ れ、ヴェラフロンス コアウイレンシス(Velafrons coahuilensis)と命 名された。ホロタイプ標本は、ほぼ幼体の完全頭骨と頸椎などの部分的 な骨格である。額には、骨質のトサカがあった。ヴェラフロンスは、コ リトサウルスやヒパクロサウルスの幼体の標本にとてもよく似ている。 頭骨は、他の属の同じ成長段階のものより大きい。通常は二足歩行、食 事の時などは四足歩行であった。体長は、約7.6m、最大で10mほどだ ったと推定されている。 |
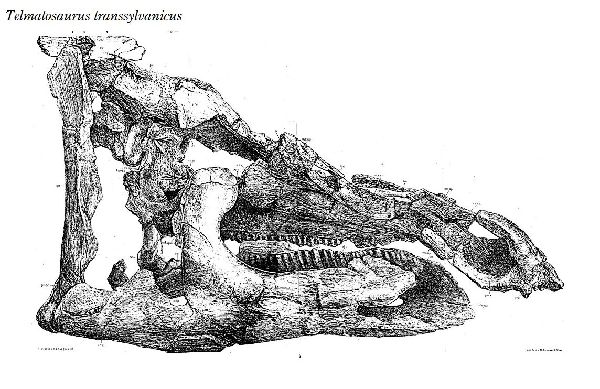
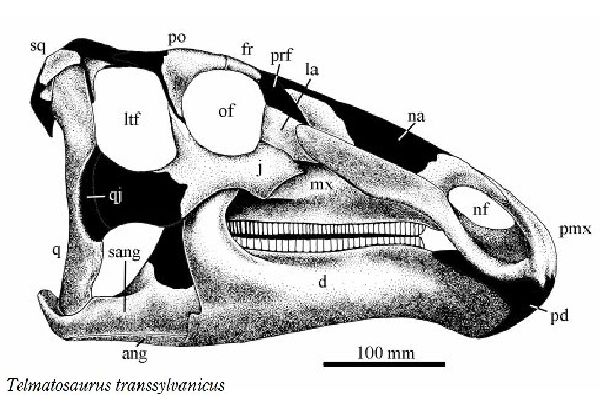
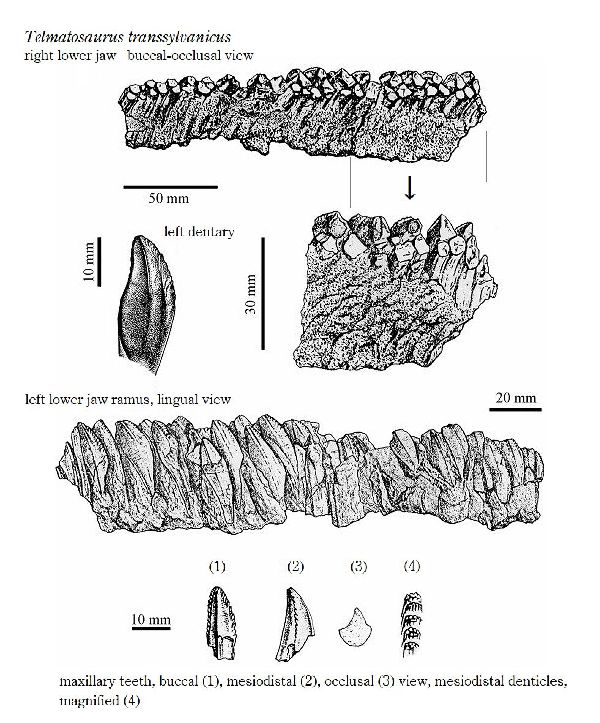
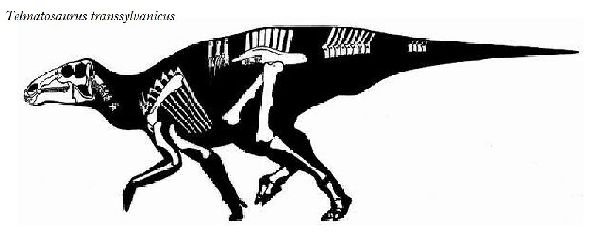
◎ テルマトサウルス (Telmatosaurus)
 |
|---|
 |
 |
 |
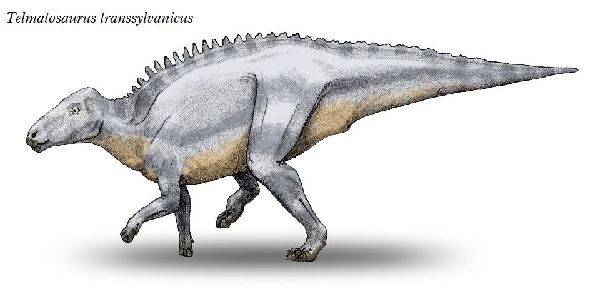 |
| テルマトサウルスは、1895年に数人の農民が、領主の娘であるイロナ・ ノプシャに彼らがトランシルヴァニアのフネドアラ地区(当時のフニャド 郡)のサチュルにあるノプシャ家の所有地で発見した恐竜の頭骨を贈呈し た。1899年、兄のフランツ・ノプシャは、この頭骨を(Limnosaurus transsylvanicus)と命名した。属名は、ギリシャ語で「沼地」を意味 する limnè からの派生で、ハドロサウルス類が沼地に生息していたと いう仮説にちなんだものである。1903年に属名がワニに使われていた ので、テルマトサウルス(Telmatosaurus)と改名した。化石は、ルーマ ニアのハツェグ盆地の白亜紀後期(6800万年前)のスンペトル層(堆積当 時は、ヨーロッパ多島海の島の一つ)から産出した。ホロタイプ標本は、 頭骨と下顎の骨で構成されている。テルマトサウルスは、基底ハドロサ ウルス科恐竜の属である。サウロロフス亜科、ランベオサウルス亜科の いずれにも属さない可能性がある。通常は二足歩行、食事の時などは四 足歩行であった。テルマトサウルスは、比較的小型のハドロサウルス類 である。ホロタイプは、幼体の標本と考えられている。体長は、約5m、 体重は、600Kgほどだったと推定されている。島に生息したため、多く の食物を食べる必要がないように小さい体になったと考えられ、大きさ については、島嶼矮化の例として説明されている。 |
ハドロサウルス類の化石紹介
鳥脚類へ
恐竜類へ
パラエオアート館へ
トップページへ